
Prenylaatiotie taas on maailmassa yleisesti lääkkein vaikutettuna, koska sen tien päädyssä on koplesterolisynteesi, jota koetetaan säätää. Mitä lääkesäätö vaikuttaa prenylaation normaalikarttaan kehossa, on toinen asia. Prenylaatiotie on kompromittoitunut syövissä.
https://media.springernature.com/lw685/springer-static/image/art%3A10.1007%2Fs12035-013-8627-z/MediaObjects/12035_2013_8627_Fig1_HTML.gif
{kind=link}
https://what-when-how.com/molecular-biology/prenylation-molecular-biology/
Prenylation (Molecular Biology)
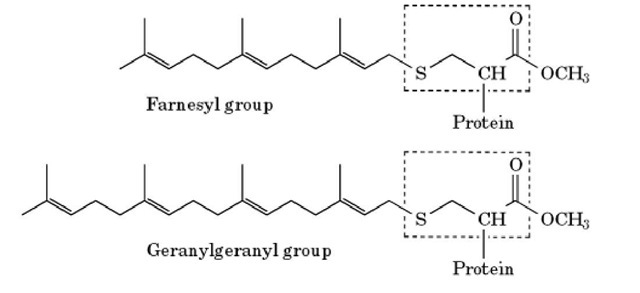
Figure 1. Modification of C-terminal cysteine residues by prenyl groups. The C-terminal cysteine residue of the protein is outlined by the dotted line. The thiol group is thioether-linked to either a farnesyl or a geranylgeranyl group, and the exposed carboxyl group is methylated.
http://what-when-how.com/wp-content/uploads/2011/05/tmp1C12_thumb.jpg
{kind=link}
Table 1. Examples of Prenylated Proteins
Farnesylated
Ras proteins
Transducin g subunit
Rhodopsin kinase
Nuclear lamins A and B
Fungal mating pheromonesa
Geranylgeranylated
g subunits of heterotrimeric G-proteins
Ras-related G-proteins (Rho/Rac/Rap/Ral/Rab)
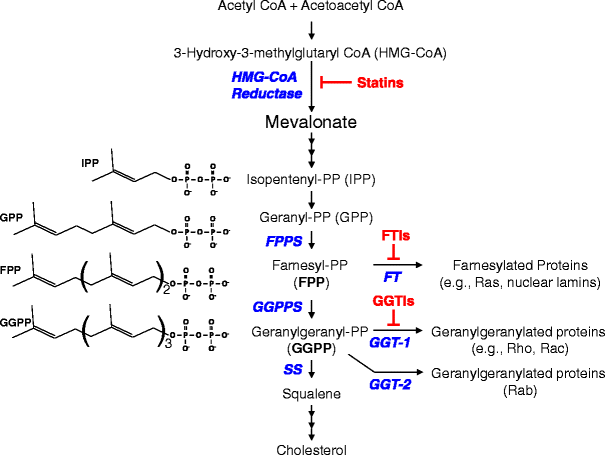
Isoprenoids are branched unsaturated hydrocarbons that are synthesized in eukaryotic cells from acetyl Coenzyme A (Acetyl CoA) by the first part of the metabolic pathway that is used to synthesize cholesterol and other sterols.
Attachment of isoprenoids to proteins is a post-translational process with four main steps:
1) recognition of the C-terminal sequence (CAAX) by one of three distinct prenyltransferases (1);
2) prenylation of a cysteine (C) residue(s) located at or close to the C-terminus using farnesylpyrophosphate (FPP) or geranylgeranylpyrophosphate (GGPP) as the substrate;
3) proteolysis of the C-terminal residues (-AAX) exposes the carboxyl group on the prenylated cysteine; and
4) the isoprenylated cysteine is recognized by a methyltransferase, which methylates the carboxyl group using S-adenosyl methionine (SAM) as the methyl donor.
Steps 1) to 3) take place in the cytosol, whereas step 4) occurs on the cytoplasmic surface of the endoplasmic reticulum (ER) or the plasma membrane (PM) .
Thus efficient methylation requires prior isoprenylation to localize the protein at the membrane surface. The thioether linkage between the cysteine and the prenyl group is chemically very stable and probably not subject to metabolic turnover. However, the carboxylic ester linkage to the methyl group is relatively labile, and may be removed after attachment. These steps differ substantially between proteins, depending on the sequence motif at the C-terminus:
1. Cys-a-a-X (CAAX) If X is serine (S) , methionine (M), or glutamine (E), it is recognized by farnesyl transferase (FTase), and the cysteine residue will be farnesylated.
If X is leucine (L), it is recognized by geranylgeranyltransferase I (GGTase-1), and the cysteine residue will be geranylgeranylated. The identity of the "a" residues (usually aliphatic) is less important, but can influence whether isoprenylation takes place or not.
Farnesyl transferase and geranylgeranyltransferase I are both heterodimers; they have identical a subunits, whereas the a subunits have only 30% identify.
Farnesylation can also occur at the C-terminus of a variety of fungal mating pheromone peptides, and in yeast the same enzyme is used for farnesylating both proteins and peptides. Although farnesyl groups have relatively low affinity for membranes themselves, they can enhance the membrane association due to other lipid groups. Farnesyl groups, because of their small size, may also play an important role in protein-protein interactions by binding directly to specific sites on other proteins (2, 3).
2. Cys-Cys, Cys-X-Cys or Cys-Cys-X-X.
(CC, CXC or CCXX) These double cysteine motifs (CC) are restricted to the Rab subgroup of Ras-related small G-proteins. The Rab protein first forms a complex with Rab escort protein (REP1, CHM).
The Rab-REP complex is then recognized by geranylgeranyltransferase II.
After prenylation, REP remains bound to Rab until it is delivered to the membrane.
REP (CHM) probably has a dual role: recognition of Rab and masking the two geranylgeranyl groups until they can be inserted into the appropriate membrane. Both cysteines are geranylgeranylated, and consequently proteolysis cannot occur. The C-terminus is not methylated in those Rab proteins ending with the sequence Cys-Cys (4).
Many of the prenylated proteins are involved in signal transduction or vesicle traffic, and the prenyl group, by facilitating rapid and reversible binding to membranes, plays an essential role in these functions (5, 6).
The membrane affinity of the prenylated proteins can be influenced by four different mechanisms (for a general discussion of factors which can affect membrane affinity of lipid anchored proteins, see Membrane Anchors):
1. The attachment of a palmitate residue (see Palmitoylation) to a cysteine close to the C-terminus reinforces the binding (eg, as in H- or N-Ras). Palmitoylation only occurs in membranes, however, so prenylation is required for it to take place (7).
2. The presence of basic residues close to the C-terminus will result in electrostatic attraction to the negatively charged bilayer surface (as in K-Ras) and increase membrane affinity (8).
3. Methylation converts the C-terminal residue from a negatively charged, hydrophilic group to an uncharged, hydrophobic group and increases membrane affinity approximately 10-fold (5, 6)). The increase in affinity is due to the hydrophobicity of the methyl group, rather than a reduction in electrostatic repulsion, because methylation gives comparable increases in binding to uncharged membranes. Methylation can have a profound influence on the cellular distribution of farnesylated proteins, because the farnesyl group is too short to provide an effective anchor by itself. Turnover of the methyl group has also been observed, and it is possible that repeated cycles of methylation and demethylation are used to regulate protein function.
4. The membrane affinity will be reduced by soluble carrier proteins, which are able to bind to the isoprenyl group(s) and mask them from the aqueous environment. This mechanism is important for the repeated releasing and recycling of Rab proteins during membrane vesicular traffic processes (9, 10)).
Previous post: Preferential Hydration (Molecular Biology)
Inga kommentarer:
Skicka en kommentar